







MANY people have puzzled over why animal species (including us) each have their particular natural maximum lifespans; why it is they deteriorate and die when they do.
A new article by Colchero and colleagues in Nature Communications sheds some light on this issue and can help us understand why further efforts at life-prolongation for humans are becoming less effective now that so few of us die before reaching old age. Perhaps a greater appreciation of the evolution and biology of ageing will encourage us to reconsider how we approach the medical care of the elderly.
As I have written earlier (here and here), we have become so very effective at stopping children and adults from dying that less than 10% of Australians currently die before the age of 65 years. However, virtually all the remaining 90% of us die over the subsequent 35 years, with 36% of all deaths in Australia occurring in the 10 years between ages 80 and 90. Chance, the way we lived our lives, and our genes influence whether we are biologically old at 70 years or can walk up mountains in our 90s. But even then, only an infinitesimally small number of us reach 110.
We are only slowly realising that the best and most expensive technologies are unable to significantly extend, with quality, the lives of the frail elderly, and that at times our efforts shade into prolongation of dying.
[caption id="attachment_68753" align="aligncenter" width="720"]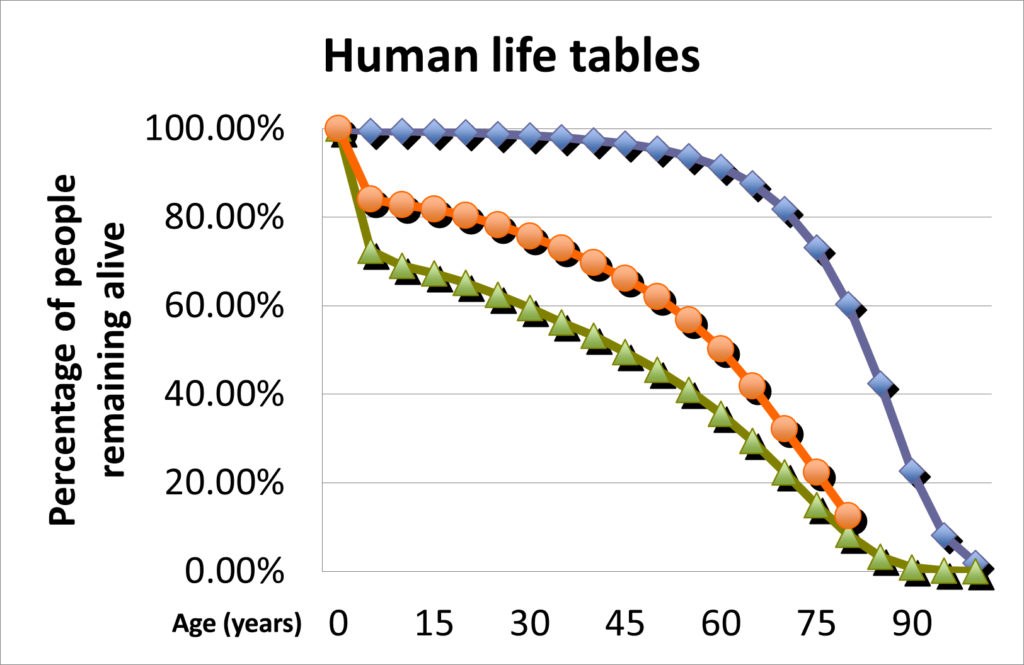 Figure 1 Human life tables: ( England and Wales 1838–1854; South Australia 1891–1900; Australia 2008–2010); maximum life-expectancy has not increased. The reduction in mortality to about 10% of the population before the age of 65 years is offset by the deaths of almost all of the remaining 90% over the subsequent 35 years. Source: Death rules[/caption]
Figure 1 Human life tables: ( England and Wales 1838–1854; South Australia 1891–1900; Australia 2008–2010); maximum life-expectancy has not increased. The reduction in mortality to about 10% of the population before the age of 65 years is offset by the deaths of almost all of the remaining 90% over the subsequent 35 years. Source: Death rules[/caption]
Colchero and colleagues collected 39 datasets on the mortality of humans and of non-human species across seven genera of other primates in a multispecies study of the relationships between the life expectancy at birth and “lifespan equality” – the latter being the spread of the distribution of the ages of deaths in a population, where equality increases with the proportion of individuals that are able to approach the species maximum possible lifespan.
The non-human primate data came from long term studies of both wild and zoo-bred populations, the latter being well nourished, free from predation and afforded veterinary care.
In summary, the study found that:
Old age itself is perhaps best described as that stage of life when the deterioration of the physical condition of adult individuals accelerates because we have exhausted the intrinsic defenses against degeneration that had evolved to keep us alive long enough to achieve sufficient survival benefit for our group.
Holding the second law of thermodynamics at bay is hard work – the disorder of the universe, its entropy, increases continuously. Individual lives, and communities of organisms, temporarily create order, borrowing from the universe in exchange for increased disorder elsewhere.
Our dynamic complexity (and that of all other organisms) does not come with any permanence in the structures that we grow on our way to adulthood. We do not have built-in repair mechanisms, redundancy or robustness to maintain our youthfulness indefinitely. Over time – at all levels from molecular to cellular and anatomical, DNA and collagen to neurones, kidneys and joints – we wear out and break down.
As individuals, we only have to live for long enough to achieve succession through our offspring. Each starts life as the order and information of a single cell, accumulating their own personal debt to disorder as their community of cells multiplies and organises, all the while sustained by the order has been borrowed by their parents and the rest of their community.
Many of those wondering about the evolutionary determinants of life-expectancy will have realised that natural selection favours investing energy in mechanisms that maintain fitness when they offer an overall species’ survival gain. This means ensuring that sufficient numbers of adults survive long enough to sustain continuity by providing essential physical and cultural support to at least one and sometimes several generations of progeny (here and here). Equally, however, no advantage has been found by natural selection from investing in mechanisms for delaying ageing beyond the time when older individuals can be succeeded in those vital roles – in our evolutionary past, hanging around after having ceased to be of survival benefit to successive generations may have even diminished the overall viability of a social species.
The timing of biological senescence co-evolved with a range of interacting and interdependent traits; gestation time, risks of childbirth, childhood mortality, age of first reproduction, time it takes to grow a complex brain, fertility, mortality in adulthood, time taken to learn vital skills such as food gathering and environmental awareness, and the need for social engagement in adulthood for success as social animals.
While the co-evolution of multiple codependent traits is clearly very complex, our natural maximum lifespan is the product of the relentless trial and error of evolution that continuously sorts out the balance across the large number of factors that shape the way we experience our sociable lives.
Over the past few hundred years, we have limited many of the hazards that shaped our evolution. Most of us now survive to become elderly because public health measures, along with modern health care technology, help us to avoid dying in childhood and adulthood. However, increasing lifespan equality cannot translate into prolonged maximum life expectancy because the biology of the timing and processes of our ageing is intrinsic to our bodies and has evolved as part of the success of complex life.
None of this should be taken as suggesting that our lives as older persons are not valuable, or that throughout our history, as older persons we have not or do not make significant contributions to the success of our families and communities. Nor that we should not be doing our best to ensure that in our old age we are secure, respected, calm and well supported, particularly when we are repaying the final installments of our debt to entropy through the disruptive pathophysiology of our ageing.
Put simply, death in old age is a complex and evolved biological reality and an integral part of what we are. And while the timing of our individual biological decline varies, almost none of us are alive after the age of 110 and, only one person is known to have lived beyond the age of 120.
Our increased “lifespan equality”, a product of our ingenuity, means that on average we each now live twice as many years as we did just 200 years ago. We have yet to address the fact that this is a major factor in the dramatic growth of the human population and the degradation of the hugely complex global environment on which we all depend.
At the level of the lives of our individual patients, it is of no credit to our profession that we have failed to fully engage with the reality of the biology of mortality in old age. Too often, as patients we are treated in the same paradigm when we become frail in old age as we were when we were younger, with the result that many of us suffer from the imposition of futile (and expensive) technology on to our deaths. This reality is being brought into sharp relief by the contrast between the challenge of finding sufficient resources to provide appropriate care for ageing persons, while nary an eyelid is batted at the application of expensive treatment that is of little benefit, or sometimes harmful.
Dr Will Cairns is Consultant Emeritus Palliative Medicine at Townsville University Hospital, and Associate Professor at James Cook University.
The statements or opinions expressed in this article reflect the views of the authors and do not represent the official policy of the AMA, the MJA or InSight+ unless so stated.
A new article by Colchero and colleagues in Nature Communications sheds some light on this issue and can help us understand why further efforts at life-prolongation for humans are becoming less effective now that so few of us die before reaching old age. Perhaps a greater appreciation of the evolution and biology of ageing will encourage us to reconsider how we approach the medical care of the elderly.
As I have written earlier (here and here), we have become so very effective at stopping children and adults from dying that less than 10% of Australians currently die before the age of 65 years. However, virtually all the remaining 90% of us die over the subsequent 35 years, with 36% of all deaths in Australia occurring in the 10 years between ages 80 and 90. Chance, the way we lived our lives, and our genes influence whether we are biologically old at 70 years or can walk up mountains in our 90s. But even then, only an infinitesimally small number of us reach 110.
We are only slowly realising that the best and most expensive technologies are unable to significantly extend, with quality, the lives of the frail elderly, and that at times our efforts shade into prolongation of dying.
[caption id="attachment_68753" align="aligncenter" width="720"]
Figure 1 Human life tables: ( England and Wales 1838–1854; South Australia 1891–1900; Australia 2008–2010); maximum life-expectancy has not increased. The reduction in mortality to about 10% of the population before the age of 65 years is offset by the deaths of almost all of the remaining 90% over the subsequent 35 years. Source: Death rules[/caption]Colchero and colleagues collected 39 datasets on the mortality of humans and of non-human species across seven genera of other primates in a multispecies study of the relationships between the life expectancy at birth and “lifespan equality” – the latter being the spread of the distribution of the ages of deaths in a population, where equality increases with the proportion of individuals that are able to approach the species maximum possible lifespan.
Understanding the nature and extent of biological constraints on the rate of ageing and other aspects of age-specific mortality patterns is critical for identifying possible targets of intervention to extend human lifespans, and for understanding the evolutionary forces that have shaped lifespans within and across speciesThe nine human databases were from populations whose mortality had not been significantly influenced by modern medicine and public health measures.
The non-human primate data came from long term studies of both wild and zoo-bred populations, the latter being well nourished, free from predation and afforded veterinary care.
In summary, the study found that:
… consistent with theory and with the long-understood effect of averting early deaths, observed variation in life expectancy and lifespan equality within each primate genus appears to be largely accounted for by variation in the pattern of early deaths, and very little by actuarial senescence.And:
Our findings support the idea that, in historical population when life expectancies were low, mortality improvements for infants, and in age-independent mortality, were the central contributors to the decades-long trend towards longer human life expectancies and greater lifespan equality. These improvements were largely the result of environmental influences including social, economic, and public health advances.This study of a group of highly socially related species supports the concept that the timing and inexorability of ageing (and hence maximum life-expectancy) is an evolved consequence of natural selection not having found benefit in living beyond a certain age. In summary, the historical pattern of our primate lives has been characterised by high infant and childhood mortality, lower death rates in adulthood, and rounded out by the re-emergence of high mortality in those who have survived to reach old age. But what is old age?
… improvements in the environment are unlikely to translate into a substantial reduction in the rate of ageing … or in the dramatic increase in lifespan that would result from such a change. It remains to be seen if future advances in medicine can overcome the biological constraints that we have identified here, and achieve what evolution has not.
Old age itself is perhaps best described as that stage of life when the deterioration of the physical condition of adult individuals accelerates because we have exhausted the intrinsic defenses against degeneration that had evolved to keep us alive long enough to achieve sufficient survival benefit for our group.
Holding the second law of thermodynamics at bay is hard work – the disorder of the universe, its entropy, increases continuously. Individual lives, and communities of organisms, temporarily create order, borrowing from the universe in exchange for increased disorder elsewhere.
Our dynamic complexity (and that of all other organisms) does not come with any permanence in the structures that we grow on our way to adulthood. We do not have built-in repair mechanisms, redundancy or robustness to maintain our youthfulness indefinitely. Over time – at all levels from molecular to cellular and anatomical, DNA and collagen to neurones, kidneys and joints – we wear out and break down.
As individuals, we only have to live for long enough to achieve succession through our offspring. Each starts life as the order and information of a single cell, accumulating their own personal debt to disorder as their community of cells multiplies and organises, all the while sustained by the order has been borrowed by their parents and the rest of their community.
Many of those wondering about the evolutionary determinants of life-expectancy will have realised that natural selection favours investing energy in mechanisms that maintain fitness when they offer an overall species’ survival gain. This means ensuring that sufficient numbers of adults survive long enough to sustain continuity by providing essential physical and cultural support to at least one and sometimes several generations of progeny (here and here). Equally, however, no advantage has been found by natural selection from investing in mechanisms for delaying ageing beyond the time when older individuals can be succeeded in those vital roles – in our evolutionary past, hanging around after having ceased to be of survival benefit to successive generations may have even diminished the overall viability of a social species.
The timing of biological senescence co-evolved with a range of interacting and interdependent traits; gestation time, risks of childbirth, childhood mortality, age of first reproduction, time it takes to grow a complex brain, fertility, mortality in adulthood, time taken to learn vital skills such as food gathering and environmental awareness, and the need for social engagement in adulthood for success as social animals.
While the co-evolution of multiple codependent traits is clearly very complex, our natural maximum lifespan is the product of the relentless trial and error of evolution that continuously sorts out the balance across the large number of factors that shape the way we experience our sociable lives.
Over the past few hundred years, we have limited many of the hazards that shaped our evolution. Most of us now survive to become elderly because public health measures, along with modern health care technology, help us to avoid dying in childhood and adulthood. However, increasing lifespan equality cannot translate into prolonged maximum life expectancy because the biology of the timing and processes of our ageing is intrinsic to our bodies and has evolved as part of the success of complex life.
None of this should be taken as suggesting that our lives as older persons are not valuable, or that throughout our history, as older persons we have not or do not make significant contributions to the success of our families and communities. Nor that we should not be doing our best to ensure that in our old age we are secure, respected, calm and well supported, particularly when we are repaying the final installments of our debt to entropy through the disruptive pathophysiology of our ageing.
Put simply, death in old age is a complex and evolved biological reality and an integral part of what we are. And while the timing of our individual biological decline varies, almost none of us are alive after the age of 110 and, only one person is known to have lived beyond the age of 120.
Our increased “lifespan equality”, a product of our ingenuity, means that on average we each now live twice as many years as we did just 200 years ago. We have yet to address the fact that this is a major factor in the dramatic growth of the human population and the degradation of the hugely complex global environment on which we all depend.
At the level of the lives of our individual patients, it is of no credit to our profession that we have failed to fully engage with the reality of the biology of mortality in old age. Too often, as patients we are treated in the same paradigm when we become frail in old age as we were when we were younger, with the result that many of us suffer from the imposition of futile (and expensive) technology on to our deaths. This reality is being brought into sharp relief by the contrast between the challenge of finding sufficient resources to provide appropriate care for ageing persons, while nary an eyelid is batted at the application of expensive treatment that is of little benefit, or sometimes harmful.
Dr Will Cairns is Consultant Emeritus Palliative Medicine at Townsville University Hospital, and Associate Professor at James Cook University.
The statements or opinions expressed in this article reflect the views of the authors and do not represent the official policy of the AMA, the MJA or InSight+ unless so stated.
Loading comments…
Newsletters
Subscribe to the InSight+ newsletter
Immediate and free access to the latest articles
No spam, you can unsubscribe anytime you want.
By providing your information, you agree to our Access Terms and our Privacy Policy. This site is protected by reCAPTCHA and the Google Privacy Policy and Terms of Service apply.